Pansen
Der Pansen (lat. pantex, über frz. panse „Wanst“; anatomisch Rumen) ist ein Hohlorgan bei Wiederkäuern (Ruminantia) und der größte der drei Vormägen. Er ist eine große Gärkammer, welche dem eigentlichen Drüsenmagen (bei Wiederkäuern als Labmagen bezeichnet) vorgeschaltet ist. Im Pansen erfolgt der Aufschluss der Zellulose durch Mikroorganismen („Pansenflora“) und die Resorption der dabei entstehenden Verbindungen. Zusammen mit dem Netzmagen (Reticulum) sorgt er für die Einleitung der Rejektion (Hochwürgen in die Mundhöhle) grober Futterbestandteile zum Wiederkäuen bzw. dem Weitertransport zerkleinerter und vorverdauter Nahrungsteile in den Blättermagen. Pansen und Netzmagen werden daher funktionell auch zum Ruminoreticulum zusammengefasst und gehen beim Embryo aus einer gemeinsamen Anlage hervor.

Magen eines Kalbs: m – Ende der Speiseröhre, v – Pansen, n – Netzmagen, b – Blättermagen, l – Labmagen, t – Beginn des Dünndarms
Inhaltsverzeichnis
1 Anatomie
1.1 Größe und Lage
1.2 Untergliederung
1.3 Gefäße und Nerven
2 Feinbau
2.1 Pansenzotten
2.2 Schleimhaut
2.3 Muskelschicht
3 Entwicklungsgeschichte
4 Funktion
4.1 Pansenflora und -fauna
4.2 Fermentation
4.3 Resorptionsvorgänge
4.4 Pansenmotorik
5 Untersuchung
6 Erkrankungen
7 Mensch und Pansen
8 Pansen als Tiernahrung
9 Umweltbelastung
10 Literatur
11 Weblinks
12 Einzelnachweise
Anatomie |
Größe und Lage |
Der Pansen ist bei ausgewachsenen Tieren der größte der drei Vormägen. Er nimmt die gesamte linke Hälfte der Bauchhöhle ein, im hinteren Bereich beansprucht er teilweise sogar einen Teil der rechten Bauchhälfte. Nur der Netzmagen liegt zwerchfellseitig auf der linken Seite noch vor dem Pansen, die Leber wird durch diese beiden Vormägen bei Wiederkäuern vollständig auf die rechte Bauchseite verdrängt. Der Pansen liegt mit seiner linken Wandfläche (Facies parietalis) direkt der Innenseite der linken Bauchwand an und reicht von der 8. Rippe bis zum Beckeneingang. Seine Eingeweidefläche (Facies visceralis) grenzt vorn an Blätter- und Labmagen, weiter hinten an das Darmkonvolut. Der Pansen hat beim erwachsenen Hausrind ein Fassungsvermögen von bis zu 100 Litern, beim Hausschaf von etwa 10 Litern und nimmt damit etwa 75 bis 80 % des gesamten Magenvolumens ein.
Untergliederung |

Mägen eines Schafes von links, 1–13 Pansen. 1 Schleudermagen, 2 dorsaler Pansensack, 3 ventraler Pansensack, 4 Recessus ruminis, 5 kaudodorsaler Blindsack, 6 kaudoventraler Blindsack, 7 Sulcus cranialis, 8 linke Längsfurche, 9 Sulcus coronarius dorsalis, 10 Sulcus coronarius ventralis, 11 Sulcus caudalis, 12 Sulcus accessorius sinister, 13 Panseninsel, 14 Pansen-Netzmagen-Furche, 15 Netzmagen, 16 Labmagen, 17 Speiseröhre, 18 Milz.
Der Pansen ist durch von außen sichtbare Furchen in mehrere Teilkammern unterteilt. Im Bereich dieser Furchen liegen im Inneren erhabene Wülste, die Pansenpfeiler (Pilae).
Die linke und rechte Längsfurche (Sulcus longitudinalis sinister und dexter) bzw. die entsprechenden Pfeiler im Inneren (Pila longitudinalis sinister und dexter) unterteilen den Pansen in seine zwei Hauptabteilungen, den rückenseitigen (Saccus dorsalis) und den bauchseitigen Pansensack (Saccus ventralis). Der vordere Teil des rückenseitigen Pansensacks überragt den bauchseitigen. Dieser Teil wird als Pansenvorhof (Atrium ruminis) oder „Schleudermagen“ bezeichnet. Mit ihm ist die Milz verwachsen. Der vordere Teil des bauchseitigen Pansensacks wird Recessus ruminis („Pansenaussackung“) genannt. Zwischen diesen beiden vorderen Abschnitten liegen Sulcus bzw. Pila cranialis. Von den linken und rechten Längsfurche gibt es eine sich abspaltende, weiter oben verlaufende zusätzliche Furche (Sulcus accessorius dexter und sinister) bzw. entsprechende Pfeiler (Pila accessoria dextra und sinistra), die die Panseninsel (Insula ruminis) abgrenzen.
Nach hinten werden die beiden Pansensäcke durch die rücken- bzw. bauchseitige Kranzfurche (Sulcus coronarius ventralis bzw. Sulcus coronarius dorsalis), im Inneren entsprechend durch die Pila coronaria ventralis bzw. dorsalis, von den beiden Pansenblindsäcken (Sacci caeci) abgegrenzt. Zwischen dem nach hinten und oben gerichteten Saccus caecus caudodorsalis und dem nach hinten und unten gerichteten Saccus caecus caudoventralis liegen Sulcus bzw. Pila caudalis. Bei den Hirschen sind drei Blindsäcke ausgebildet.
Vom nach vorn angrenzenden Netzmagen (Reticulum) wird der Schleudermagen durch die Pansen-Netzmagen-Furche (Sulcus ruminoreticularis) abgegrenzt. Nur für den Sulcus ruminoreticularis gibt es keinen entsprechenden Pfeiler im Pansen. Stattdessen findet man hier eine Plica ruminoreticularis. In diesem Grenzbereich zwischen beiden Vormägen mündet von oben die Speiseröhre.
Der Pansen ist nur in einem kleinen Bereich des vorderen rückenseitigen Pansensacks mit der Bauchwand verwachsen und wird durch die Schwerkraft, seinen Inhalt und die übrigen inneren Organe in seiner Lage gehalten. An den Längsfurchen heftet sich das große Netz (Omentum majus) an. Dabei verläuft dessen tiefe Wand (Paries profundus) an die rechte Längsfurche, umschließt den Pansen und setzt sich an der linken Längsfurche als oberflächliche Wand (Paries superficialis) fort. Der Paries superficialis verläuft wieder bauchwärts um den Pansen herum und zieht zur rechten oberen Rumpfwand sowie zum Blätter- und Labmagen.
Gefäße und Nerven |
Die Blutversorgung erfolgt durch die rechte und linke Pansenarterie (Arteria ruminalis dextra und sinistra), die aus der Milzarterie (Arteria lienalis) entspringen. Die rechte Pansenarterie ist die größere der beiden Pansenarterien. Sie zieht in der rechten Längsfurche nach hinten, schlägt im Sulcus caudalis auf die Bauchwandseite des Pansens um und versorgt damit auch den hinteren Teil der linksseitigen Pansenwand. Die linke Pansenarterie zieht über den Sulcus cranialis ebenfalls auf die Bauchwandseite. Die entsprechenden Venen ergießen sich in die Pfortader (Vena portae) und sorgen so dafür, dass die im Pansen aufgenommenen Nährstoffe direkt in die Leber gelangen.
Die Lymphgefäße ziehen zu mehreren Lymphknotengruppen der Magenlymphknoten (Lymphonodi gastrici). Die rechten Pansenlymphknoten (Lymphonodi ruminales dextri) liegen entlang der rechten, die linken (Lymphonodi ruminales sinistri) in der linken Pansenlängsfurche. Zudem sind die vorderen Pansenlymphknoten (Lymphonodi ruminales craniales) im Sulcus cranialis und die Pansen-Labmagen-Lymphknoten (Lymphonodi ruminoabomasiales) an der vorderen Unterseite des Pansen im Bereich der Berührungsfläche zum Labmagen in die Lymphdrainage einbezogen.
Die nervale Steuerung (Innervation) des Pansens erfolgt durch das vegetative (autonome) Nervensystem. Der parasympathische Nervus vagus (X. Hirnnerv) zieht in Form eines oberen und unteren Stammes (Truncus vagalis dorsalis und ventralis) zusammen mit der Speiseröhre an den Pansen. Die Hauptversorgung erfolgt durch den oberen Stamm, der untere beteiligt sich nur an der Innervation des Schleudermagens. Die efferenten (zum Pansen hinziehende) Nervenfasern der Vagusstämme regen die Motorik des Pansens an, die afferenten (vom Pansen wegziehende) leiten Impulse von Mechano- und Chemorezeptoren in das verlängerte Mark (Medulla oblongata). In diesem Teil des Stammhirns liegt auch das Reflexzentrum, welches die Pansenbewegungen ohne Beteiligung des Bewusstseins steuert. Die sympathischen Nervenfasern gelangen über das Bauchhöhlengeflecht (Plexus coeliacus) mit den Blutgefäßen zum Pansen. Ihre Efferenzen wirken hemmend auf die Vormagenbewegungen, die Afferenzen leiten Schmerzreize. Die Kontraktion der glatten Muskulatur des Pansens wird über die Ganglienzellen des darmeigenen Nervensystems vermittelt, geordnete Bewegungen (siehe Pansenmotorik) sind jedoch ohne den Vaguseinfluss nicht möglich. Die Nervenzellen des darmeigenen Nervensystems liegen zwischen den beiden Muskelschichten des Pansens in Form des Plexus myentericus (Auerbach-Plexus). Der Plexus submucosus (Meissner-Plexus) ist, im Gegensatz zu den übrigen Abschnitten des Magen-Darm-Kanals, an den Vormägen nicht ausgebildet.
Feinbau |

Histologisches Präparat eines Pansens vom Schaf. 1 Pansenzotten, 2 Epithel, 3 Schleimhaut, 4 Verbindungsschicht, 5 Muskelschicht, 6 Bauchfell

Schleimhaut des Pansens eines Schafes mit Pansenzotten; in der Mitte ein Pansenpfeiler
Der Pansen besteht wie alle inneren Hohlorgane aus einer innenliegenden Schleimhaut, einer Muskelschicht aus glatter Muskulatur und dem außen anliegenden Bauchfell. Im Bereich der dem Pansen aufliegenden Milz verwächst der obere vordere Teil des Pansens mit der Bauchwand, so dass ein kleines Gebiet ohne Bauchfellüberzug ist.
Pansenzotten |
Die Schleimhaut bildet, im Gegensatz zum ersten Kompartiment der Vormägen der Kamele, bei Wiederkäuern Pansenzotten (Papillae ruminis) zur Oberflächenvergrößerung, beim Rind etwa um den Faktor 7. Die Größe, Form und Verteilung dieser Zotten variiert nach der Ernährungsweise und in Abhängigkeit von der aktuell verfügbaren Nahrung. Die Zotten entstehen bereits embryonal und sind fadenförmig mit rundem bis ovalen Querschnitt, bei erwachsenen Tieren sind sie meist zungenförmig, variieren aber nach Nahrungsangebot (s. u.). Die bei großen Wiederkäuern etwa 300.000 größeren Zotten („Hauptzotten“) sind beim Rind bis zu 13 mm, bei Giraffen bis zu 25 mm lang.
Bei den sogenannten Gras- und Raufutterfressern (z. B. Rinder, Schafe, Mufflon) sind die Pansenpfeiler meist zottenfrei und Pansendach und -boden besitzen nur wenige, kurze Zotten. Bei den „Selektierern“, also jenen Wiederkäuern die Rohfaser-arme, leichtverdauliche Pflanzen aufnehmen (z. B. Reh, Elch, Giraffe), sind die Zotten eher gleichmäßig verteilt und meist auch auf den Pfeilern ausgebildet. Beim „Intermediärtyp“, also jenen Tieren die Pflanzen beider Kategorien aufnehmen (z. B. Rothirsch, Ziegen, Impala), sind die Zotten im Bereich der Pfeiler nur kurz.
In Abhängigkeit vom Nahrungsangebot und damit der Zusammensetzung der bei der Gärung entstehenden Fettsäuren kommt es zu Zottenveränderungen. So nehmen unter kargen Ernährungsbedingungen (Winter, Trockenzeit) die Zotten in Zahl, Länge und Dicke stark ab („Hungerzotten“) und gleichen dann eher den fadenförmigen Zotten der Föten. Dieser Vorgang ist umkehrbar (reversibel), der Anpassungsvorgang dauert etwa zwei bis drei Wochen. Am stärksten ausgeprägt sind die Veränderungen beim Intermediärtyp. Die Anpassungsvorgänge betreffen nicht nur die Zotten, sondern auch die Eigenschicht der Schleimhaut und die Blutgefäßarchitektur.
Schleimhaut |
Die Schleimhaut des Pansens ist drüsenlos und trägt ein mehrschichtiges verhorntes Plattenepithel.
Das Pansenepithel ist im Hinblick auf Resorptionsfunktion des Pansens modifiziert und unterliegt zyklischen, durch Resorptionsvorgänge bedingten Veränderungen. Das Epithel wird in fünf Schichten untergliedert, die bei optimalen Ernährungsbedingungen zumeist jeweils nur aus einer Zelllage bestehen. Neben den eigentlichen Epithelzellen kommen auch durch das Epithel wandernde weiße Blutkörperchen und Langerhans-Zellen vor, die der Immunabwehr dienen.

Lichtmikroskopisches Bild einer Pansenzotte. 1 Ballonzelle, 2 Körnerzelle, 3 Parabasalzelle, 4 Basalzelle, 5 Eigenschicht, 6 Papillarkörper, 7 Zottenzentralarterie
- Das Stratum basale des Epithels liegt der Basalmembran an. Die hochprismatischen Basalzellen sind über Hemidesmosomen an der Basalmembran verankert und besitzen einen relativ großen Zellkern. Die Zwischenräume zwischen den Basalzellen sind relativ weit, die Verbindung der Zellen untereinander erfolgt durch auf langen Zytoplasmafortsätzen sitzende Desmosomen.
- Die tiefe Stachelzellschicht (Stratum spinosum profundum) des Epithels besteht aus den polygonalen Parabasalzellen. Sie haben ebenfalls weite, durch Zellfortsätze überbrückte Zwischenzellräume. Die Zellfortsätze reichen teilweise ebenfalls bis zur Basalmembran. Die Zellkontakte verhalten sich wie bei den Basalzellen.
- Die oberflächliche Stachelzellschicht (Stratum spinosum superficiale) setzt sich aus den abgeflachten, oberflächenparallel angeordneten Intermediärzellen zusammen. Ihre Fortsätze sind besonders reich an Tonofilamenten und Haftplatten zur Bildung von Desmosomen. Die Intermediärzellen enthalten viele Lysosomen.
- Die Körnerzellschicht (Stratum granulosum) des Epithels besteht aus zwei Typen von Körnerzellen. Die Typ A-Körnerzellen ähneln noch den Intermediärzellen, haben aber nur noch kurze Zellfortsätze. Sie enthalten kleine Zellkerne und kleine Keratohyalin-Granula. Die Typ B-Körnerzellen sind größer und haben bereits stark geschrumpfte (pyknotische) Zellkerne. Sie bilden keine zusammenhängende Zellschicht und enthalten schollige Ansammlungen von Keratohyalin. Durch Verschmelzung der Zellmembranen kommt es in den zum Innenraum gerichteten Körnerzellen zur Ausbildung von fester Zellverbindungen (tight junctions).
- Die zum Hohlraum des Pansens liegende Hornschicht (Stratum corneum) besteht aus Hornzellen. Die Typ A-Hornzellen sind bereits abgeplattet und mit Keratohyalin-Schollen und Zellkernresten ausgefüllt. Sie sind über tight junctions untereinander verbunden. Aus ihnen entstehen die Typ B-Hornzellen mit schwammartig aufgelockertem Keratin. Diese Zellen können sich aus dem Epithelverband lösen oder sich weiter in die Typ C-Hornzellen (Ballon- oder Quellzellen) entwickeln. Letztere sind durch weitere Schwellung ballonförmig aufgetrieben, die Zellorganellen sind nahezu verschwunden und die gewellte Zellmembran kann schließlich einreißen. Ballonzellen fehlen meist in den zottenfreien Regionen und bei schlechtem Nahrungsangebot.
Die Eigenschicht der Schleimhaut (Lamina propria mucosae) bildet einen Papillarkörper. Die Eigenschicht besteht aus kollagenem und elastischem Bindegewebe mit einigen Abwehrzellen (Leukozyten, Plasmazellen, Mastzellen). In ihr liegen die feinen Blutgefäße und Nervenfasern. Dabei ziehen aus den Gefäßen der Eigenschicht an der Basis der Pansenzotten zwei Zottenrandarterien und eine oder zwei Zottenzentralarterien in die Zotten, von denen parallel zum Epithel verlaufende Arteriolen ausgehen. Diese speisen ein dichtes Kapillarnetz direkt unter dem Epithel. Die abführenden Venolen liegen ebenfalls direkt unter dem Epithel und haben ein Endothel mit Poren zur Erleichterung des Stoffaustauschs. Die Venolen sind zum Teil zu Sinusoiden erweitert. Eine Schleimhautmuskelschicht (Lamina muscularis mucosae) fehlt zwar, aber das Bindegewebe der Eigenschicht ist zu einer Lamina compacta verdichtet, die auch einige glatte Muskelzellen enthält. Die Eigenschicht geht fließend in die Verbindungsschicht (Tela submucosa) mit ihren Gefäßnetzen über. Lymphfollikel und Drüsen sind in der Schleimhaut des Pansens nicht ausgebildet.
Muskelschicht |
Wie der gesamte Magen-Darm-Trakt besteht die Muskelschicht aus einer inneren Ring- und einer äußeren Längsmuskelschicht, diese Anordnung ist allerdings modifiziert. Die Längsmuskelschicht des übrigen Magen-Darm-Kanals strahlt nur in den rückenseitigen Pansen- und Pansenblindsack ein, sie entspricht den äußeren schiefen Fasern (Fibrae obliquae externae) des einhöhligen Magens. Die Ringmuskelschicht erstreckt sich dagegen nur auf den Schleudermagen und die bauchseitigen Pansenabschnitte. Zu dieser jeweiligen Muskellage kommt eine zweite Muskelschicht, die auch die Muskelschleife am Mageneingang bildet und den inneren schiefen Fasern (Fibrae obliquae internae) des einhöhligen Magens entspricht. Sie umfasst ringförmig alle Pansenabschnitte. Durch diese zusätzliche Schicht besteht auch die Muskelschicht des Pansens an allen Stellen aus zwei Lagen mit unterschiedlicher Verlaufsrichtung.
Entwicklungsgeschichte |
Während sich die Trennung der Huftiere in Paar- und Unpaarhufer bereits im frühen Eozän vollzog, begann die Evolution der Wiederkäuer und damit des Pansens erst im Oligozän und hatte ihren Höhepunkt im Miozän. Dabei entwickelte sich bei den Stammformen der Wiederkäuer und den Kamelen ein mehrkammriger Magen, der zunächst nur der Kurzvergärung diente, ähnlich den rezenten Selektierern (wie Rehe). Erst mit der Entwicklung der Verzögerungsmechanismen im Vormagensystem (Netzmagen-Blättermagen-Öffnung) konnten auch schwerverdauliche und rohfaserreiche Futterpflanzen verwertet werden.
Im Gegensatz zu den Wiederkäuern besteht der Magen der Kamele aus drei drüsenhaltigen Kompartimenten. Gelegentlich wird auch das erste Kompartiment der Kamele als „Pansen“ bezeichnet, was aufgrund der morphologischen Unterschiede jedoch vermieden werden sollte. Solche Vormägen mit einem mehrschichtig verhornten Plattenepithel besitzen auch Faultiere, Nabelschweine und Wale (mit Ausnahme der Schnabelwale). Bei Kloakentieren und Schuppentieren ist der gesamte Magen so ausgekleidet.
Beim Wiederkäuer-Embryo entstehen alle Magenabteilungen aus der spindelförmigen Magenanlage, wie sie auch bei den anderen Säugetieren zu finden ist. Die Magenanlage dreht sich bei Wiederkäuern bei der ersten Magendrehung nur um 90° nach links. Der Pansen entsteht zusammen mit dem Netzmagen (Haube) im Bereich der großen Krümmung (Curvatura major) als eine links-vorn-oben gerichtete Ausbuchtung, die Pansen-Hauben-Anlage. Bereits beim 20 mm langen Rinderembryo sind alle vier Magenabschnitte angelegt. Mit dem Größenwachstum kommt es auch zum Auswachsen der beiden Blindsäcke, die zunächst nach vorn-oben gerichtet sind. Mit der Rückbildung der Urniere vollzieht der Pansen eine Drehung über die Rückenseite, so dass die Blindsäcke nach hinten verlagert werden und der Pansen seine definitive Position einnimmt.
Zur Geburt ist der Pansen mit einem Volumenanteil von 47 % noch die größte Magenabteilung, dicht gefolgt vom Labmagen (etwa 40 %). Während der Säugezeit in den ersten Lebenswochen übertrifft der Labmagen durch stärkeres Wachstum den Pansen an Größe, da die Muttermilch dort verdaut wird und über den Haubenrinnenreflex an den Vormägen vorbeigeleitet wird. Erst mit der Aufnahme strukturwirksamer Rohfaser entwickelt sich der Pansen unter dem Einfluss mechanischer und chemischer Reize (durch die Fermentation entstehende Fettsäuren) nun zur deutlich größten Magenabteilung. Dabei kommt es auch zur oben beschriebenen Umwandlung der Pansenzotten. Die Besiedlung des Pansens mit Mikroorganismen erfolgt beim Jungtier vor allem durch Kontakt mit anderen Tieren, Bakterien werden auch mit dem Futter aufgenommen.
Funktion |
Im Pansen liegt eine Schichtung des Inhalts vor. Im unteren Abschnitt befindet sich die flüssige Phase. Darauf schwimmen im mittleren Abschnitt gröbere Pflanzenbestandteile und oben entsteht eine Gasblase. Die Flüssigphase mit ihren festen Schwebeteilchen und Mikroorganismen wird als Pansensaft bezeichnet.
Pansenflora und -fauna |
Die Gesamtheit der Mikroorganismen im Pansen wird als Pansenflora und -fauna bezeichnet. Es handelt sich um vorwiegend anaerobe (nur unter Sauerstoffabschluss lebensfähige) Bakterien, Einzeller (sogenannte „Infusorien“) und Pilze. Sie machen etwa 20 % des Volumens des Panseninhalts aus.
Die Bakterien spalten Kohlenhydrate (Zellulose, Hemizellulose, Pektine, Xylane, Zucker) und Proteine. Im Pansen sind etwa 1010 bis 1011 Bakterien/ml vorhanden, die vorwiegend an den Oberflächen der Nahrungspartikel und des Pansenepithels anhaften. Sie gehören zu etwa 200 verschiedenen Arten, unter anderem Ruminococcus spp., Lactobacillus spp., Clostridium spp. und Bacteroides spp. Neben den Abbauprozessen sind die Bakterien auch an der Aufrechterhaltung des Pansenmilieus beteiligt. Die am Epithel haftenden sauerstoffverzehrenden Bakterien halten das anaerobe Milieu aufrecht und das negative Redoxpotential von −250 bis −300 mV sorgt dafür, dass der Abbau der Kohlenhydrate nur bis zu den kurzkettigen Fettsäuren und nicht vollständig zu Kohlendioxid und Wasser erfolgt. Die im Pansensaft befindlichen Archaeen bilden aus Kohlendioxid und Wasserstoff Methan und senken somit den Wasserstoff-Partialdruck im Pansen, was eine übermäßige Bildung von Ethanol und Milchsäure verhindert. Methan ist für den Wiederkäuer nicht verwertbar und muss zusammen mit dem Kohlendioxid als „Abgas“ über den Ruktus abgegeben werden. Die Abgabe dieses Kohlenwasserstoffs senkt jedoch die Effizienz der Energieverwertung.
Die Protozoen bilden etwa die Hälfte der Biomasse der Pansenflora und setzen sich vor allem aus Wimpertierchen (105 bis 108/ml, vor allem Vertreter der Isotrichidae und Ophryoscolecidae) und in geringerem Maß aus Geißeltierchen (103 bis 104/ml) zusammen. Protozoen sind in geringerem Umfang am Kohlenhydrat- und Eiweißabbau (ca. 10 %) beteiligt. Sie können leicht abbaubare Kohlenhydrate aufnehmen und verhindern so deren überstürzten Abbau und damit eine Pansenazidose infolge zu hoher Mengen an organischen Säuren. Außerdem können die Protozoen schädliche Futterbestandteile (toxische Pflanzeninhaltsstoffe und Schwermetalle) abbauen oder binden. Darüber hinaus nehmen die Protozoen Bakterien auf und regulieren damit deren Population. Die Protozoen scheinen jedoch für die Vormagentätigkeit nicht unbedingt notwendig zu sein, vielfache Studien belegen gar, dass sie für eine ineffiziente Stickstoffnutzung verantwortlich sind. Die Effizienz der Stickstoffnutzung kann durch ein Entfernen der Protozoen aus dem Pansen, der sogenannten Defaunierung, gesteigert werden. Zudem kann so die Methanproduktion verringert werden, da Protozoen von Methanbildnern als Wirt genutzt werden.[1]
Über die Bedeutung der im Pansen vorkommenden Pilze (Neocallimastigaceae) liegen bislang noch keine gesicherten Erkenntnisse vor. Sie verwerten in geringem Ausmaß lösliche Kohlenhydrate und Proteine und sind auch zur Bildung langkettiger Fettsäuren befähigt. Ihr Vorkommen gilt ebenfalls als nicht zwingend erforderlich.
Fermentation |
Von der Pansenflora werden die β-glykosidischen Bindungen von Strukturkohlenhydraten aufgebrochen, vor allem die von Zellulose, die von den Verdauungsenzymen der Säugetiere nicht gespalten werden kann. Der so entstehende Traubenzucker (Glucose) dient den Mikroorganismen als Substrat, die Produkte ihres Stoffwechsels sind unter anderem kurzkettige Carbonsäuren wie zum Beispiel Propionsäure, Buttersäure und vor allem Essigsäure. Zum Wachstum benötigen die Mikroorganismen neben fermentierbaren Kohlenhydraten auch Stickstoff, der durch das im Futter enthaltene Protein, aber auch durch Nicht-Protein-Stickstoff (NPN) geliefert wird. Proteine werden durch die Mikroorganismen im Pansen weitgehend zu Peptiden, Aminosäuren oder Ammoniak gespalten und dienen ihnen anschließend als Stickstoffquelle.
Der Wiederkäuer stellt also den Mikroorganismen die Fermentationskammer und das Substrat zur Verfügung. Die Mikroorganismen liefern dem Wiederkäuer:
Energie: Ein wesentliches Produkt der Mikroorganismen sind flüchtige Carbonsäurederivate. Bei einem Rind entstehen etwa 5 mol kurzkettige Fettsäuren je kg Trockensubstanzaufnahme, bei einer Hochleistungsmilchkuh allein etwa 45 mol Azetat pro Tag.[2] Diese werden durch die Pansenwand vom Wiederkäuer resorbiert.
Proteine: Die Mikroorganismen selbst verbleiben nicht dauerhaft im Pansen, sondern verlassen diesen nach und nach, während durch Vermehrung neue entstehen. Im Dünndarm werden die Mikroorganismen selbst dann vom Wiederkäuer weitgehend verdaut und liefern dem Tier dadurch Proteine. Daher sind die Mikroorganismen selbst eine wichtige Proteinquelle für die Wiederkäuer und ermöglichen es ihnen damit indirekt, auch anorganische Stickstoffquellen zu verwerten.
Vitamine u. a.: Neben Energie und Protein liefern die Mikroorganismen dem Wiederkäuer auch verschiedene Vitamine u. a.; beispielsweise können die Mikroorganismen Cobalamin (Vitamin B12) und viele andere synthetisieren.
Zur Aufrechterhaltung des für Mikroorganismen erforderlichen Milieus sind verschiedene Mechanismen vorhanden, beispielsweise wirken die flüchtigen Fettsäuren im Pansen pH-senkend. Der physiologische pH-Wert des Pansens beträgt 5,5 bis 7. Bei niedrigeren pH-Werten (Pansenazidose) wird das Milieu für die Mikroorganismen ungünstig. Dem wird u. a. durch den beim Wiederkauen mit abgeschlucktem Speichel entgegengewirkt, da hier puffernde Substanzen (v. a. Bikarbonat, HCO3 und Hydrogenphosphat, HPO42−) enthalten sind. Je nach Futteraufnahme des Wiederkäuers (Hochleistungskühe: >25 kg Trockenmasse) können pro Tag bis zu 270 Liter Speichel gebildet werden.[2]
Resorptionsvorgänge |
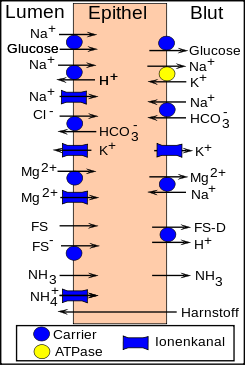
Resorptionsvorgänge im Pansen (FS=Fettsäuren, FS−=dissoziierte FS, FS-D=Fettsäurederivate)
Das Pansenepithel stellt eine Barriere gegen passive Resorptionsvorgänge dar, es hält chemische Gradienten zwischen Panseninhalt und Blut aufrecht und wird daher als „mäßig dichtes Epithel“ bezeichnet. Diese Barrierefunktion verhindert ein Übersäuern des Blutes. Für die im Pansen stattfindenden umfangreichen Resorptionsvorgänge gibt es verschiedene zelluläre Transportproteine für Mineralstoffe (Natrium, Chlorid, Kalium, Magnesium, Calcium). Insbesondere die Magnesiumresorption spielt eine große Rolle, da sie beim Wiederkäuer praktisch ausschließlich im Pansen stattfindet und ein Magnesiummangel bei Rindern im Frühjahr nicht selten ist (Weidetetanie). Phosphat wird vermutlich nur passiv und in geringer Menge über Diffusion zwischen den Pansenepithelzellen (parazellulär) aufgenommen. Wasser wird unter Normalbedingungen nur in geringer Menge aufgrund osmotischer Gradienten resorbiert.
Die bei der Gärung entstehenden kurzkettigen Fettsäuren werden vermutlich passiv über einen Stoffgradienten aufgenommen, sowohl dissoziiert als auch undissoziiert. Dabei werden diese zu einem großen Teil in den Pansenepithelzellen chemisch umgesetzt (Buttersäure in Ketokörper, Propionsäure in Milchsäure) ehe sie an das Blut abgegeben werden. Dadurch bleibt der Gradient zwischen Lumen und Epithelzelle erhalten und die Epithelzelle nutzt diese Stoffe zur Energiegewinnung, die für die aktiven Transportprozesse notwendig ist.
Das beim Proteinabbau entstehende Ammoniak liegt bei physiologischen pH-Werten größtenteils als Ammonium-Ion (NH4+) vor und wird größtenteils durch die Pansenflora zur Proteinsynthese verwendet und zum Teil auch über Kaliumkanäle in die Epithelzellen aufgenommen. Bei hohen pH-Werten (Pansenalkalose) entsteht mehr Ammoniak, welches aufgrund seiner Lipidlöslichkeit leicht in die Zellen eintreten kann und von der Leber entgiftet werden muss. Verschiedene Hersteller bieten deshalb Futtermittel mit pansenbeständigem Protein an. Die beim Eiweißabbau entstehenden Aminosäuren und Peptide werden im Pansen nicht oder nur in Spuren resorbiert.
Harnstoff kann über den Speichel sowie über Blut und Pansenepithel in das Panseninnere abgegeben werden und steht so der mikrobiellen Proteinsynthese zur Verfügung. Im Gegensatz zu anderen Säugetieren, wo der Harnstoff über den Urin ausgeschieden werden muss, sind Wiederkäuer in der Lage, diesen wiederzuverwerten, bei eiweißarmer Fütterung bis zu 90 %. Diesen Vorgang bezeichnet man als Pansen-Leber-Kreislauf (ruminohepatischer Kreislauf). Ein Teil des Harnstoffs wird über die Milch abgegeben (→ Milchharnstoff).
Neben Harnstoff können auch Kalium, H+, HCO3− und Wasser in den Pansen zurücktransportiert werden.
Pansenmotorik |
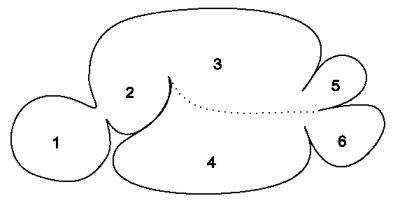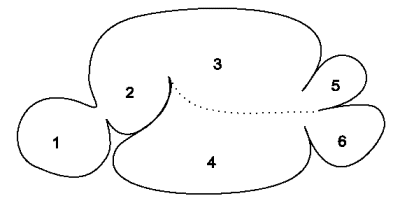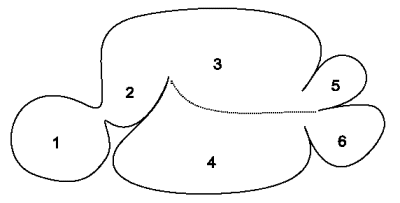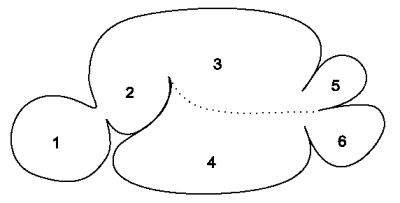
Der mikrobielle Aufschluss, das Wiederkäuen und der Weitertransport werden durch eine komplizierte Abfolge von Muskelkontraktionen, die sogenannte Pansenmotorik, aufrechterhalten.
Dabei sorgen diese Kontraktionen der Pansenmuskulatur und der Pansenpfeiler für eine ständige Durchmischung des Inhalts. Dabei unterscheidet man sogenannte A-Zyklen mit Beteiligung des Netzmagens und B-Zyklen, die allein im Pansen stattfinden. Die Steuerung dieser Bewegungen erfolgt über das Reflexzentrum im Stammhirn und wird afferent über die Dehnungs- und Chemorezeptoren der Pansenwand und Chemorezeptoren des Duodenums, efferent über den Nervus vagus vermittelt. Bei Fieber und Schmerzzuständen wird die Pansenmotorik gehemmt.
Beim Wiederkäuen wird durch die Kontraktion des Netz- und Schleudermagens ein Brocken grober Futterbestandteile vor den Mageneingang gehoben, durch eine Einatmung bei angehobenen Gaumensegel angesaugt und durch eine gegenläufige Kontraktionswelle (Antiperistaltik) der Speiseröhre in die Maulhöhle zurückbefördert (rejiziert). Anschließend wird die Nahrung mit den Zähnen zerkleinert und wieder in den Pansen abgeschluckt.
Die bereits feinzerkleinerten und weitestgehend zersetzten Nahrungsteile einschließlich der Mikroorganismen sammeln sich aufgrund höherer Dichte im bauchseitigen Pansensack. Über die Kontraktion dieses Pansenteils gelangen sie in den Netzmagen, der sie in den Blättermagen weitertransportiert.
Die Abgabe der sich im rückenseitigen Pansensack ansammelnden Gärungsgase erfolgt durch einen durch den Nervus vagus vermittelten Reflex, den sogenannten Ruktus. Das Gas wird durch eine Kontraktion des rückenseitigen Pansensacks während eines B-Zyklus zur sich reflektorisch öffnenden Speiseröhrenmündung transportiert und von dort über eine Antiperistaltik der Speiseröhre in Richtung Maul geleitet. Da das Gaumensegel den Weg verschließt und das Maul geschlossen ist, gelangt das Gas zunächst in die Lunge, wo das Kohlendioxid teilweise resorbiert wird, was schnelle Atembewegungen auslöst. Ein Ruktus erfolgt etwa zweimal pro Minute.
Untersuchung |
Die Untersuchung der lebensnotwendigen Pansenmotorik gehört zu jeder klinischen Allgemeinuntersuchung bei Wiederkäuern. Sie kann durch Beobachtung, Abhören oder eine in die linke Hungergrube gedrückte Faust ertastet werden. Bei einem gesunden Tier lassen sich zwei bis drei Kontraktionszyklen in zwei Minuten nachweisen. Die wichtigste funktionelle Untersuchung ist die Entnahme von Pansensaft über eine über das Maul eingeführte Pansensonde und dessen Untersuchung. Dabei werden vor allem der pH-Wert und mikroskopisch die Pansenflora beurteilt.
Erkrankungen |
Die Pansenflora befindet sich in einem empfindlichen Gleichgewicht. Bei Verschiebungen des pH-Wertes in den sauren (Pansenazidose) oder alkalischen Bereich (Pansenalkalose) kommt es zu schweren Schädigungen der Pansenflora, des Pansenepithels und zu vermehrter Bildung ungünstiger Stoffwechselprodukte (siehe den Abschnitt Funktion). Ursachen sind meist Fütterungsfehler wie zu energiereiches Futter (Getreide) mit mangelndem Rohfasergehalt (Azidose) oder energiearmes und eiweißreiches Futter (Alkalose). Futterumstellungen führen zu einer Artenverschiebung der Pansenflora und Umbauvorgängen der Schleimhaut. Abrupte Futterumstellungen (>15 % der Ration) können neben pH-Wert-Verschiebungen und Störung des Gleichgewichts der Flora auch Verhornungsstörungen des Epithels (Hyperkeratose, Parakeratose) hervorrufen. Sie müssen daher vermieden und auf 10 bis 20 Tage gestreckt werden, um den Anpassungsvorgängen den nötigen Zeitraum einzuräumen. Ein Futterentzug über wenige Tage (z. B. bei Transporten) führt gleichfalls zu schweren Störungen der Pansenflora und des Stoffwechsels des Tieres (Alkalose, Ketose). Ebenso muss bei Kälbern eine allmählich steigende Zufütterung von Grünfutter vor dem Absetzen erfolgen, um die physiologische Entwicklung des Pansens und der Pansenflora anzuregen. Bei einem Ausfall des Haubenrinnenreflexes bei Kälbern gelangt Milch in den Pansen und wird dort fehlvergoren, was Verdauungsstörungen und Durchfälle hervorruft.
Aufgrund einer Fütterung mit stärke- oder eiweißreichem Futter kann es zu schaumiger Gasblasenbildung im Pansen kommen. Es entsteht eine Pansentympanie (Aufblähung), da die Gase nicht mehr über den Ruktus abgegeben werden können. Dann muss die Gasansammlung entweder durch schaumbrechende Medikamente und einen in den Pansen eingeführten Schlauch oder in Notfällen durch einen Pansenstich mit einem Trokar beseitigt werden.
Eine Störung der Pansenmotorik kann durch Schädigung des Nervus vagus beim sogenannten Hoflund-Syndrom oder bei einem Mangel an strukturwirksamer Rohfaser auftreten. Bei einem Erliegen der Pansenmotorik (Pansenatonie) sind sowohl die mikrobiellen Abbauprozesse als auch die Gasabgabe schwer gestört, was schnell zu einem lebensbedrohlichen Zustand führt.
Eine Entzündung der Pansenschleimhaut (Ruminitis) tritt vor allem bei der Pansenazidose sowie bei Aufnahme chemisch schädigender Stoffe oder heißer Futtermittel auf. Sie kann auch durch verschiedene Bakterien (Arcanobacterium pyogenes, Fusobacterium necrophorum) bei vorheriger Schleimhautschädigung verursacht werden oder als Begleiterscheinung bei einigen Viruserkrankungen (Maul- und Klauenseuche, Bösartiges Katarrhalfieber) vorkommen.
Bei der Fremdkörpererkrankung bohren sich die Fremdkörper meist durch den Netzmagen, seltener durch den Pansen. Hier wird zumeist ein Flankenschnitt durchgeführt, die Oberfläche der Pansenwand abgetastet und anschließend über einen Pansenschnitt der Fremdkörper aus dem Netzmagen oder Pansen entfernt. In den Pansen eingelegte Magnete können einer Fremdkörpererkrankung durch metallische Gegenstände vorbeugen. Bei Verfütterung stark verschmutzten Futters kann es zu einer Versandung des Pansens kommen, die bei stärkerem Ausmaß operativ beseitigt werden muss.
Bei einer schweren Schädigung der Pansenflora, die z. B. auch durch Antibiotika oder toxische Stoffe ausgelöst werden kann, muss diese durch Übertragung von Pansensaft von einem anderen Tier wiederhergestellt werden.
Mensch und Pansen |
Als Kutteln bezeichnet man Pansenstücke zum menschlichen Verzehr, die zuvor einer aufwändigen küchentechnischen Vorbereitung unterzogen werden müssen.
| Nährstoffgehalt je 100 g grünem Pansen[3] | |||
TS | 28 g | Ca | 120 mg |
Rohfett | 5 g | P | 130 mg |
vRP | 19 g | Mg | 40 mg |
ME | 0,58 MJ | K | 100 mg |
Rohfaser | 1,1 g | Na | 50 mg |
Pansen als Tiernahrung |

Pansenpresse im Seegrenzschlachthof (1929)
Pansenstücke werden auch als Tiernahrung, vor allem für Hunde, verwendet. Sie sind in einigen kommerziellen Hundefuttermitteln enthalten und werden auch getrocknet als Pansensticks angeboten. Pansen kann auch frisch („grüner“ Pansen oder geputzt) zur individuellen Herstellung von Hundefutter genutzt werden. Aufgrund der Zusammensetzung eignet sich Pansen jedoch nicht als Alleinfuttermittel.
Umweltbelastung |
Die im Pansen entstehenden Gase sind nicht ohne Weiteres nutzbar. Die etwa täglich 200 Liter Pansengase (60 % CO2, 40 % Methan) einer Kuh stellen, insbesondere in Regionen mit hoher Rinderdichte, eine Umweltbelastung dar (siehe Treibhauseffekt). Der Pansen ist jedoch auch ein natürliches Vorbild für die Herstellung von Biogas (Methan) in technischen Anlagen. 2005 gelang es Forschern der Ohio State University, mit Bakterien der Pansenflora eine mikrobische Brennstoffzelle herzustellen.
Literatur |
Physiologie des Magen-Darm-Kanals. In: W. v. Engelhardt, G. Breves (Hrsg.): Physiologie der Haustiere. 2. Auflage. Enke-Verlag, Stuttgart 2000, ISBN 3-8304-1039-5, S. 313–422.- R. R. Hoffmann, B. Schnorr: Die funktionelle Morphologie des Wiederkäuer-Magens. Enke-Verlag, Stuttgart 1982, ISBN 3-432-88081-2.
- M. Kressin, B. Schnorr: Embryologie der Haustiere. 5. Auflage. Enke-Verlag, Stuttgart 2006, ISBN 3-8304-1061-1.
- N. Rossow: Erkrankungen der Vormägen und des Labmagens. In: N. Rossow (Hrsg.): Innere Krankheiten der landwirtschaftlichen Nutztiere. Gustav Fischer Verlag, Jena 1984, S. 224–259.
- Franz-Viktor Salomon: Magen, Ventriculus (Gaster). In: Salomon u. a. (Hrsg.): Anatomie für die Tiermedizin. 2. erw. Auflage. Enke-Verlag Stuttgart 2008, ISBN 978-3-8304-1075-1, S. 272–293.
Weblinks |
- Pansenbakterien zur Stromerzeugung
Einzelnachweise |
↑ Optimierung der mikrobiellen Umsetzungen im Pansen durch die Fütterung. (Memento vom 13. März 2014 im Internet Archive) (PDF; 845 kB).
↑ ab Jörg R. Aschenbach: Der Säure-Basen-Haushalt im Pansen. In: Vet-MedReport. V10, 2009, S. 2.
↑ J. Kamphues u. a. (Hrsg.): Supplemente zu Vorlesung und Übungen in der Tierernährung. 9. Auflage. Schaper, Hannover, S. 250.


